到19世纪50年代早期,鱼类营养研究开始不断发展,直到19世纪70年代才逐渐认识到鱼类日粮中矿物质的需求和重要性。
90种自然存在的无机矿物元素中,大约有29种(包括铜、锌、铁、锰等元素)被认为是包括鱼类在内的所有养殖动物所必需的。
因此,对微量元素进行系统的认识和研究,方能推动行业健康、高效、可持续发展。
本文作者将从以下几个方面对鱼虾微量元素铜营养进行介绍,希望能为大家认识铜营养和在饲料中选择应用铜源提供一定的参考。
一、鱼虾铜元素组织分布特点
1、鱼虾所有组织均有铜的分布:肝(胰)脏铜含量相对较高
1.1 普遍数据显示
心脏和肝(胰)脏单位重量铜含量最高(是其他脏器组织的1.5-5倍,对虾肝(胰)脏铜含量相比肌肉铜含量甚至超过10倍)。
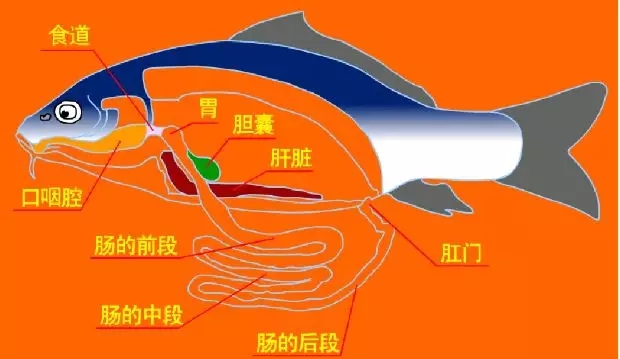
1.2 其次为
肾脏、脾脏、肠道、骨骼;
肌肉含量相对偏低。
提示:铜在肝(胰)脏和心脏等组织器官中具有相对重要的生理生化功能。
2、野生和养殖情况下微量元素铜含量存在差异
朱国萍(2010)等研究显示:野生对虾铜含量明显高于养殖对虾。
提示:可能是饲料铜营养剂型和剂量不合理,导致养殖对虾组织微量元素水平不足。
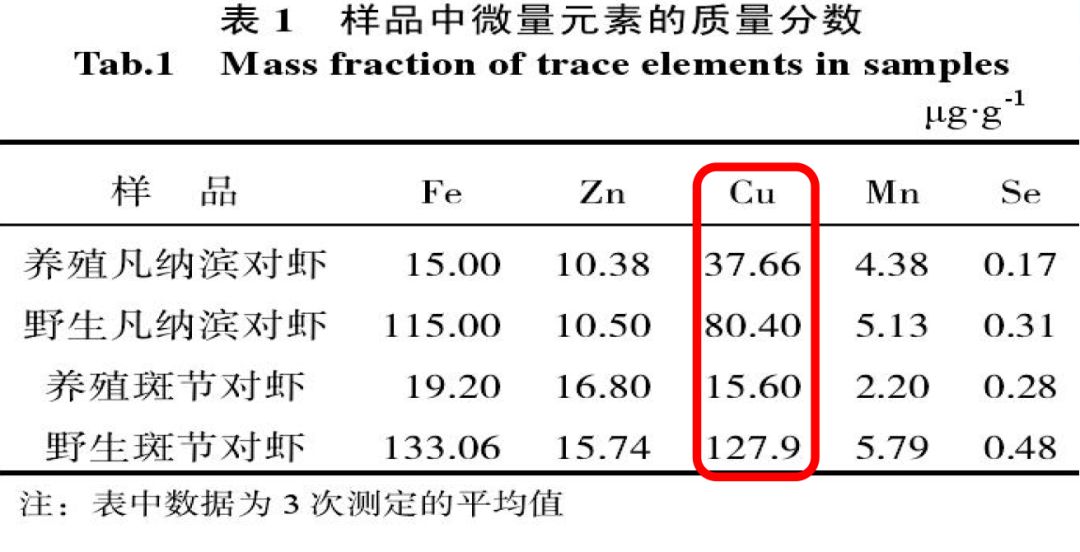
(朱国萍,2010年)
3、不同生长发育时期微量元素铜含量存在差异
费志良(2005)等研究显示:蜕皮间期肝胰脏、肌肉、卵巢铜含量显著高于软壳期,而甲壳铜含量显著低于软壳期。
提示:组织铜含量剧烈变化,说明微量元素铜在对虾蜕壳过程中发挥这重要的生理作用,补充高效的铜源更有利于对虾蜕壳的顺利进行。
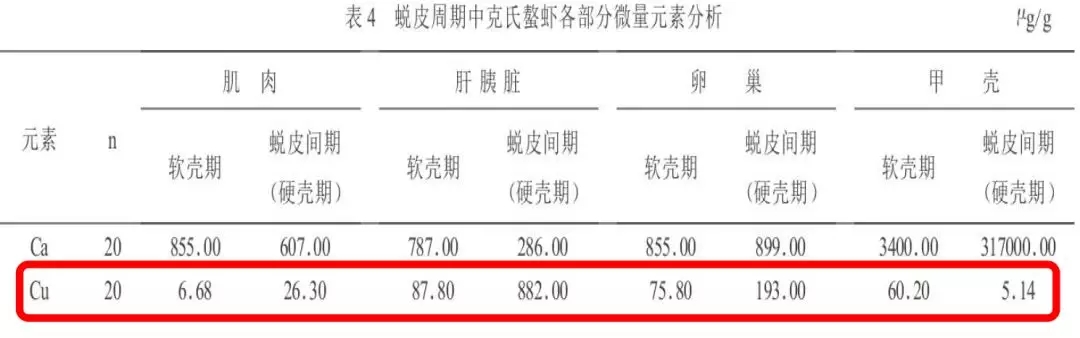
(费志良等,2005年)
赵振伦(1994)等研究显示:随鱼体生长,铜元素占比例下降。
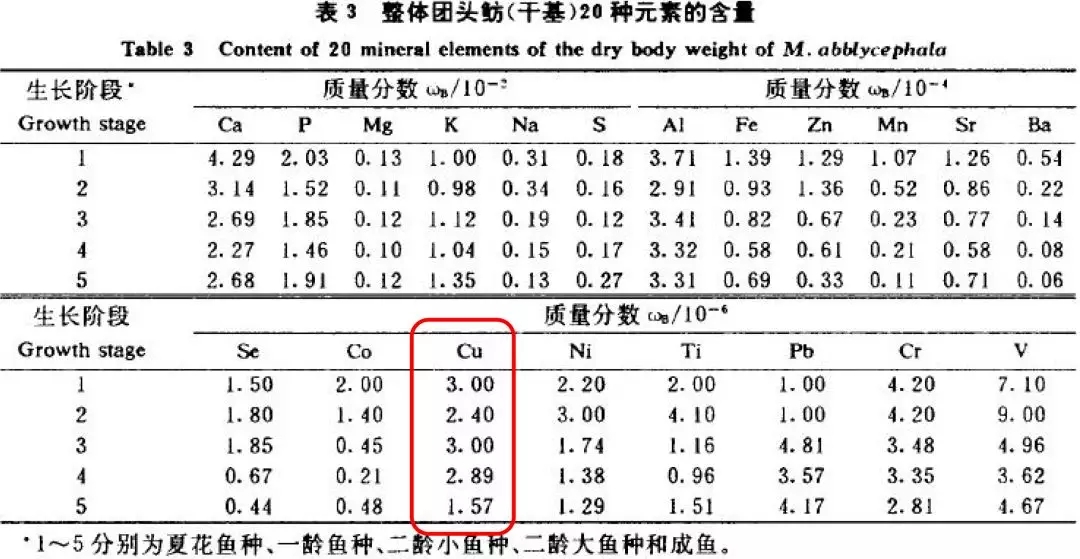
(赵振伦,1994年)
4、不同食性或种类鱼或虾微量元素铜含量差异较小
高一娜(2016)研究显示:不同食性或种类鱼微量元素差异总体无显著差异。
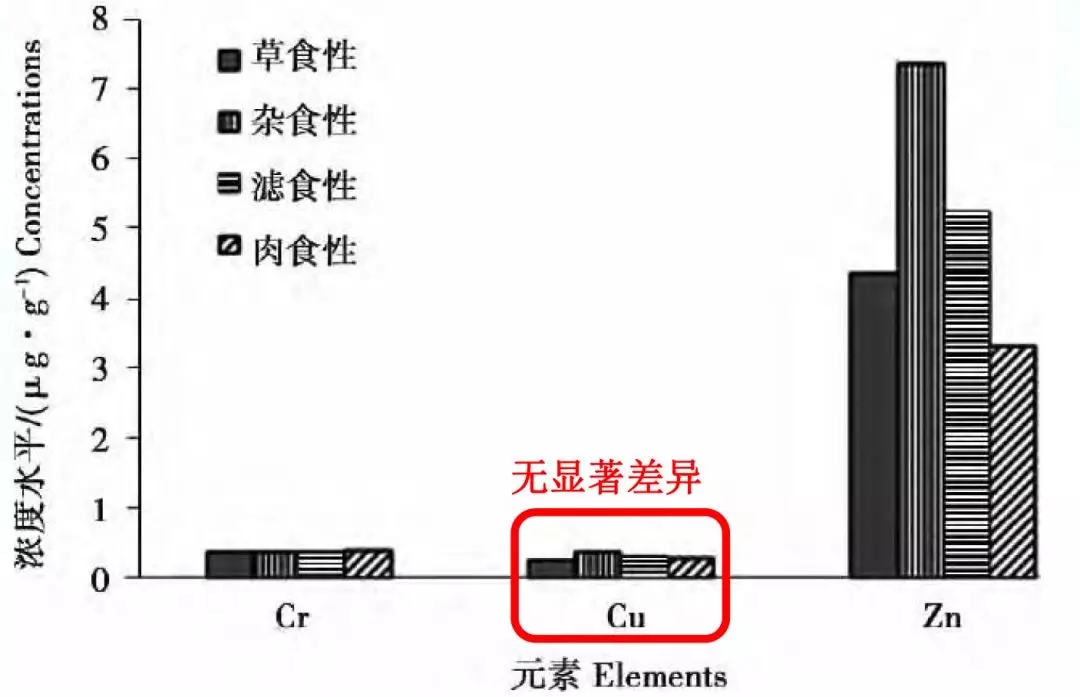
(高一娜等,2016)
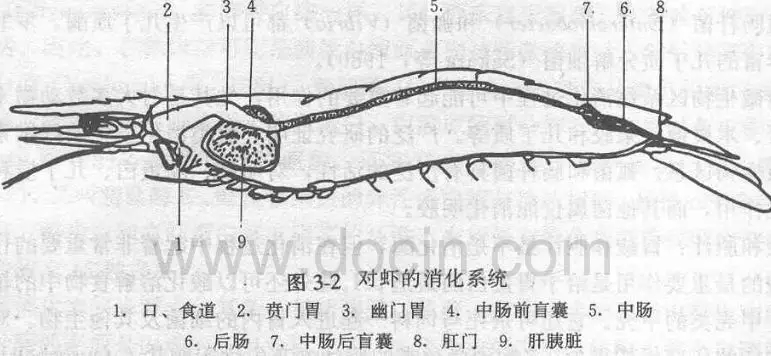
5、鱼与虾相比微量元素铜含量差异大:虾>鱼
从朱国萍(2010),高一娜(2016),王友慧、 叶元土等(2005)研究结果可以发现,对虾微量元素含量,总体上看,均明显高于鱼类。
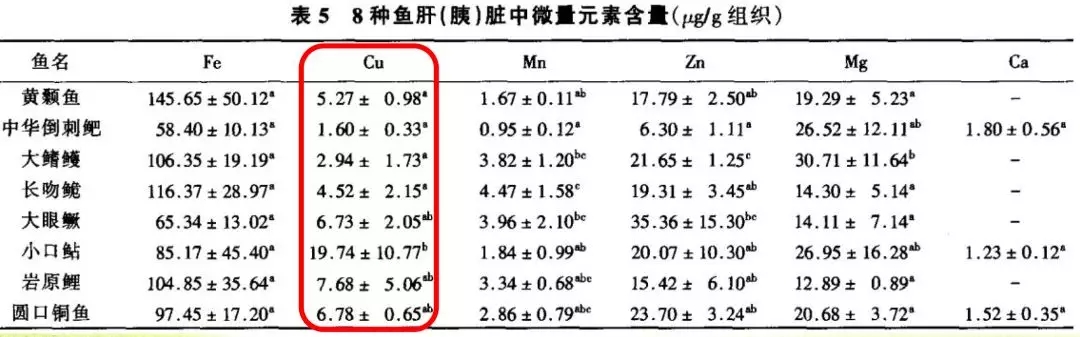
(王友慧、 叶元土等,2005)
二、鱼虾铜营养功效及作用机理
(一)鱼虾铜营养功效
铜是鱼虾必需微量元素之一,主要构成铜蓝蛋白、细胞色素C氧化酶(COX)、赖氨酰氧化酶(LOX)、CuZn-超氧化物歧化酶(SOD)、酪氨酸酶(TYR)、铁氧化酶(Ferroxidase)、多巴胺-β-羟化酶等酶的组分,这些酶与造血、机体免疫、骨骼形成、组织色素沉积等一些列重要生理生化功能有关。
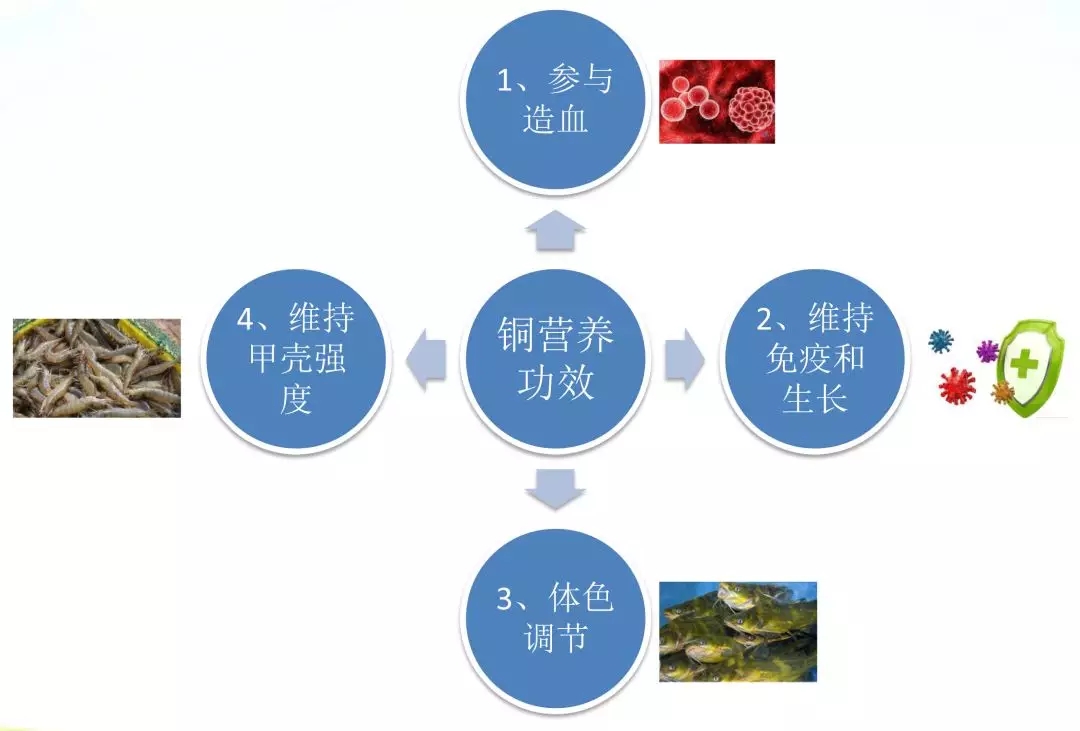
(二)作用机理
1、参与造血
1.1 鱼类血浆中90%的铜以铜蓝蛋白的形式存在
铜蓝蛋白又称铁氧化酶,铜是铜蓝蛋白的辅基, 主要参与血浆中铁的利用和提高铁传递蛋白质中铁饱和度的比率 ,在铁的转运过程中起重要作用,进而影响血红蛋白的合成。
1.2 甲壳动物血液以血蓝蛋白为氧载体,虾体内约40-50%的铜存在于血蓝蛋白中
相比鱼类,铜对虾类影响更显著。
造血机能最终影响鱼虾整体营养代谢和免疫、健康。
提示:主要与虾血浆内主要含血蓝蛋白而非血红蛋白。因此,导致实际生产中,饲料中微量元素铜的应用差异显著。
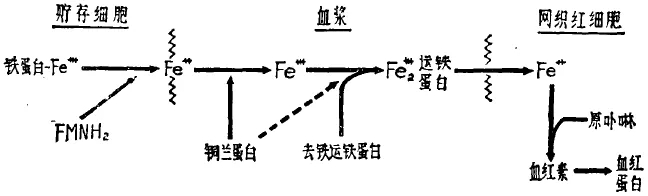
( 谈介凡 ,1984译)
2、维持免疫和生长
2.1 P. Antony Jesu Prabhu等(2014)认为矿物质代谢和免疫机制之间存在某些共同调节通路。
2.2 饲料中添加适量的铜能够提高血清铜蓝蛋白、酚氧化酶和肝胰腺总超氧化物歧化酶、铜锌超氧化物歧化酶活性。(周双艳,2014)
2.3 有研究显示:铜缺乏,免疫组织出现萎缩。
3、体色调节
![]()

图3.1 养殖武昌鱼体色变化及鳞片色素细胞的变化
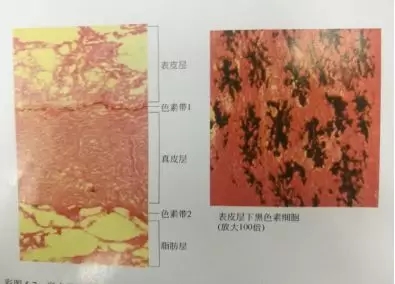
图3.2 半点叉尾鮰皮肤纵切面和表皮层基部的黑色素细胞
3.1 发生体色变化的鱼类以前主要是无鳞鱼,目前发现在有鳞鱼类,如武昌鱼、草鱼、鲤鱼等中也经常发生体色变化。体色变化的主要特征是颜色变浅( 变白) 。
3.2 铜是酪氨酸酶的辅酶成分,如果铜离子缺乏,则酪氨酸酶的活性降低,黑色素合成受到抑制,将引起水产动物体色变浅或发白。(叶元土等,2006;赵宇江,2009。)
4、维持甲壳强度
![]()

(Pechenik,2000)
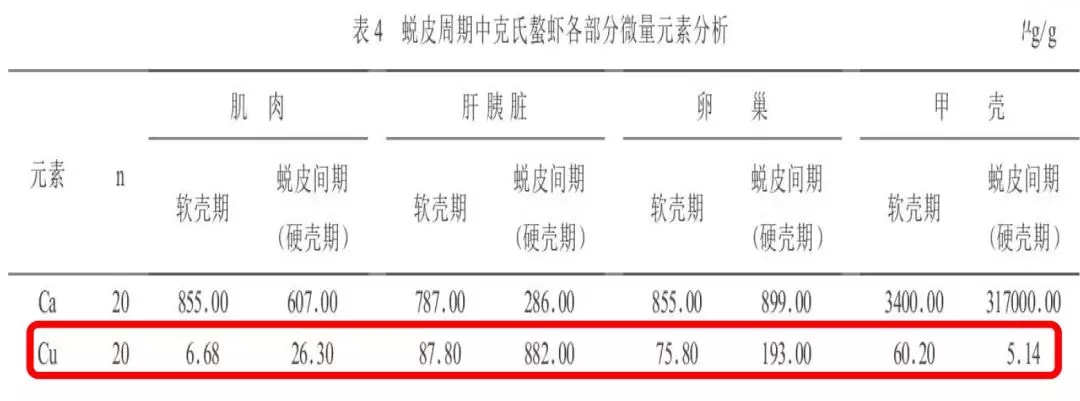
(费志良等,2005年)
4.1 铜是参与纤维化的赖氨酰氧化酶等酶的辅助因子,对维持正常骨骼强度具有重要的意义。甲壳作为虾蟹类的外骨骼,其作用与脊椎动物的骨骼有相似,因此铜对于维持甲壳强度同样具有重要意义。
4.2 蜕皮周期中虾甲壳等组织铜元素含量的剧烈变化,一定程度上反应铜在蜕壳过程中的重要作用。
当然,除以上4个重要作用外,在其它动物应用方面发现,铜对神经系统、繁殖、脂质和能量代谢、内分泌均有影响。(张春善,2007)
是否对亲鱼、亲虾、亲蟹等繁殖性能有影响?
因此,非常有必要加强对铜在水产动物方面的应用和理论研究,充分挖掘铜的营养价值,提高有限资源的使用效率和效果。
(三)铜缺乏症
铜缺乏症:含铜酶活性降低、饲料转化率降低、生长迟缓、死亡率上升。
当饲料中铜不足时,首先表现为肝脏中含铜量降低;其次表现为骨骼发育受到限制,与铜有关的酶(如细胞色素C氧化酶、铜-锌超氧化物酶等)活性下降,最终导致鱼虾生长迟缓,死亡率上升,饲料转化率降低。(艾春香,2000)南美白对虾铜缺乏,还表现为心脏增大。
三、鱼虾铜的吸收和代谢
(一)吸收位点
1、鱼虾可以通过鳃和消化道吸收铜营养
鰓主要吸收水体游离铜离子和部分颗粒吸附态铜, 通过血液循环进入肝脏;(廖伏初,2008)正常日粮浓度情况下,水体铜贡献约10%。
从食物中获得的铜进入消化道后, 在胃部就开始吸收, 但主要的吸收部位在前肠。
在普遍使用生石灰消毒的养殖水体,会使得水体游离铜离子急剧减少。所以,保证饲料足够安全有效的铜才是一种明智的选择。
(二)吸收通道和代谢途径
1、 铜的吸收主要有两大通道
1.1 离子形式,以特异或非特异性金属离子吸收通道吸收:Cu2+首先被细胞膜上的Fre1/Fre2还原酶还原为Cu+, 再通过上皮细胞的钠通道(EnaC)或者运铜载体 1(CTR1)进入细胞。(廖伏初,2008)
1.2 分子形式,以氨基酸或者小肽途径吸收假说:Cu2+与氨基酸络(螯)合,或直接补充氨基酸络合铜,到达上皮粘膜后,以氨基酸或者小肽载体,被快速吸收。
提示:氨基酸络(螯)合物具有更高效的吸收效率。
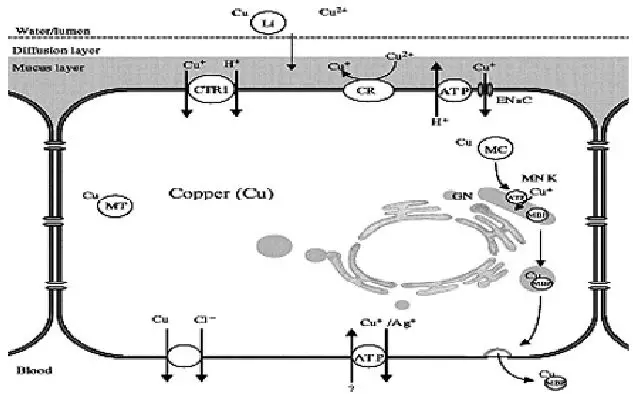
铜的代谢途径
2.1 上皮细胞吸收的铜,在胞内通过氨基酸配合物的形式,穿过肠系膜进入血液,经门静脉迅速进入肝组织,被肝细胞摄取。此外, 一些通过胆汁排出的铜,经肝肠循环也可以再回收利用。
提示:氨基酸络(螯)合物具有更高效的转运效率。
2.2 铜的排泄主要通过分泌胆汁进行, 经粪便排出。尿液中也排出少量的铜。此外,鱼的鳃也是过剩铜的排泄器官之一。
四、鱼虾铜营养需要量研究
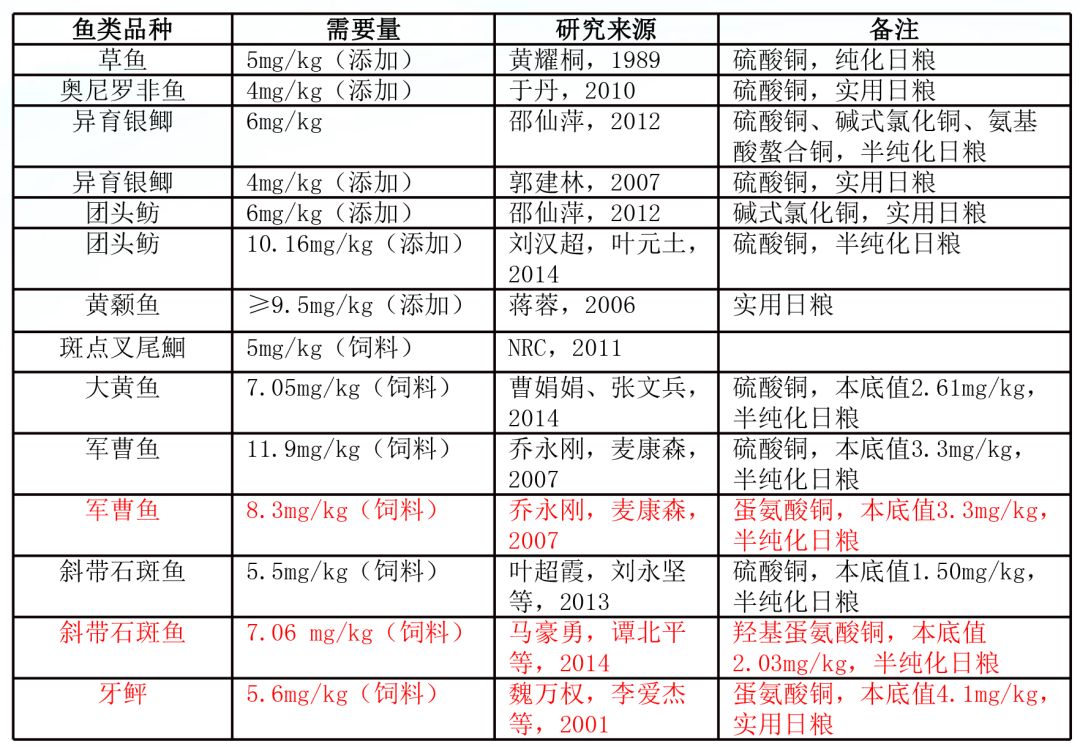
1、鱼类对铜营养的需求:从研究数据来看,正常水体微量元素铜无法满足鱼类生长需要,必需通过饲料补充。而草食性、杂食性、肉食性鱼类,对外源无机微量元素铜的需求基本一致,在4-10mg/kg之间。有机铜研究相对较少,但是已有的研究显示相对无机铜在利用率和促生长、提高免疫水平等方面具有其独特的优势。
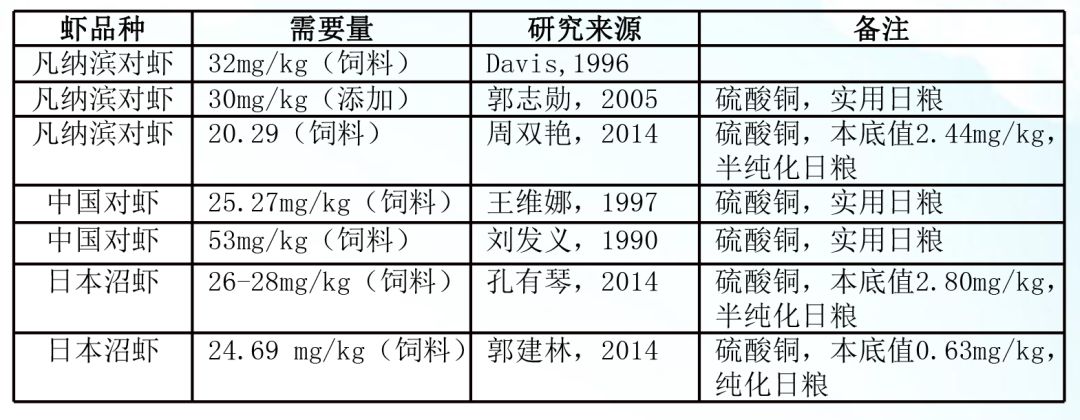
2、虾对铜营养的需求:同样正常水体微量元素铜无法满足虾类生长需要,必需通过饲料补充。不同种类虾对外源无机微量元素铜的需求基本一致,主要在20-30mg/kg之间。有机铜需求量研究相对较少,需要进一步研究。
NRC2011 鱼和虾铜推荐量分别为3和30mg/kg饲料。
微量元素铜需求普遍以增重率为评价指标,一般认为以增重为评价标准,比使用全鱼或脊椎骨微量元素浓度作为相应标准,要低估18-40%。
(P. Antony Jesu Prabhu,2014;曹娟娟,2014)
五、影响鱼虾铜吸收利用的因素
1、铜的化学形式
2、饲料中植酸、草酸、硫代葡萄糖苷及非淀粉多糖等抗营养因子
3、饲料中高钙、高铁、高锌、硫、钼拮抗因子(主要对无机盐)
4、健康状况(应激VS非应激)
5、水体铜含量
6、饲料铜含量
7、饲料动植物蛋白含量、比值和氨基酸添加水平
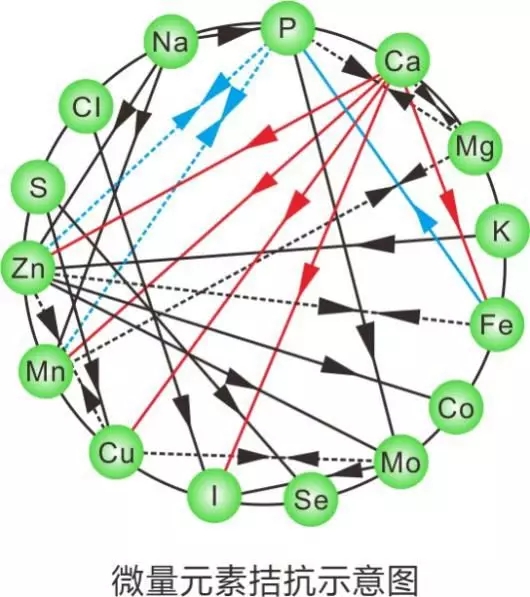
1.1 铜的化学形式有普通无机盐、有机酸盐、碱式盐、氨基酸螯合盐等,已有研究表明氨基酸络(螯)合铜比无机硫酸铜利用率要高的多。可能与有机微量元素具有更好的吸收、储存和利用特性有关。
1.2 植酸能与铜形成难溶性的盐,植物纤维也可降低铜的吸收。草酸虽然不影响铜的生物利用率,但是有协同纤维抑制铜吸收的作用。高钙等可抑制铜的吸收。鱼体生理状况的好坏很大程度决定着对Cu的吸收利用。部分游离氨基酸可以促进铜离子的吸收。
表 长江不同水系环境单元过滤水元素的背景值,ug/L
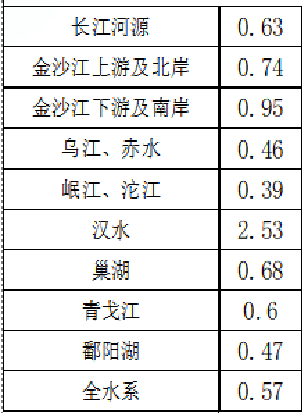
结果显示:长江水系不同环境单元河水铜含量相对变化差异较小。
表 兴嘉生物饲料原料微量元素数据库(摘录部分)
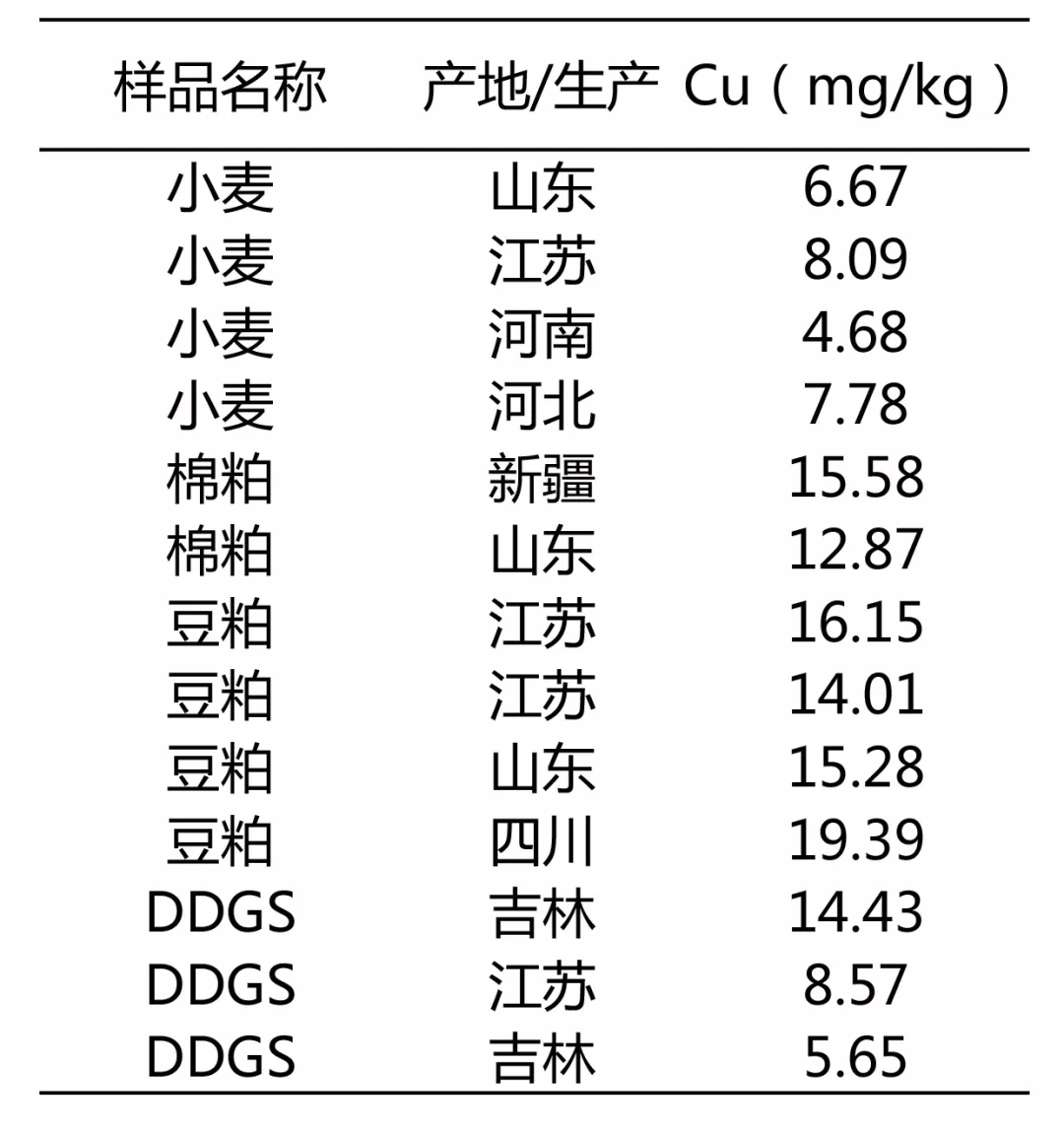
从原料大数据分析:普水鱼饲料铜本底值含量约10-15ppm,南美白对虾饲料铜本底值含量约12-20ppm。
虽然有研究显示成年动物对日粮铜的吸收率5-10%, 广泛的研究显示,水体和饲料原料微量元素含量仍无法有效满足高密度、高生产性能要求,特别是抗应激、抗病等功能营养要求下水产动物对微量元素营养的需求。
六、鱼虾铜营养选择关注点及建议
实际日粮,逐渐低蛋白化和蛋白原料多元化,使得使用硫酸铜进行添加时,拮抗铜吸收的因素增多和复杂化。使得企业在应用过程中,疑难重重,效果不稳定、不理想。
以及水产养殖环保(排放问题)和集约化、规模化快速发展,后抗生素时代的来临。企业在微量元素选择,需要考虑的维度更多。
实际生产到底如何选择微量元素
铜的营养添加剂?
(一)鱼虾铜营养选择关注点
1、微量元素铜对预混料品质的影响
铜原子半径较小,有空的电子轨道,在光线作用下Cu2+ 能于空气中的氧分子作用形成活化络合物“Cu2+....O2 ”,活化的氧分子有利于进攻维生素,脂肪等易氧化营养物质分子,使之自动氧化破坏。
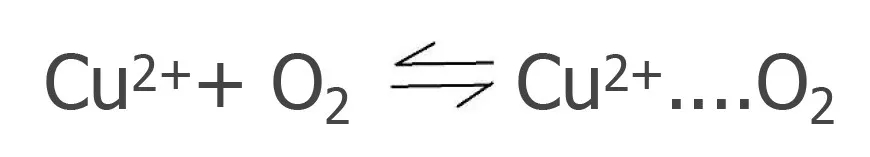
营养物质(维生素、油脂等)+ Cu2+....O2 →氧化型营养物质+H2O2 +Cu2+
更糟糕的是和亚铁离子同时存在时,亚铁离子除有类似上面催化氧化营养物质分子作用外,还能与自动氧化过程中的产物H2O2发生进一步反应,生成极为活泼的羟基自由基,而这个羟基自由基能更加强烈地氧化营养物质分子。
好的CuSO4•5H2O产品游离水<0.1%,差的产品>0.5%。CuSO4遇水易解离出Cu2+ ,因此在饲料实际生产中,特别是预混料生产中,易导致维生素、油脂等的氧化破坏。研究认为氧化性为Cu2+>Fe2+>Zn2+>Mn2+。
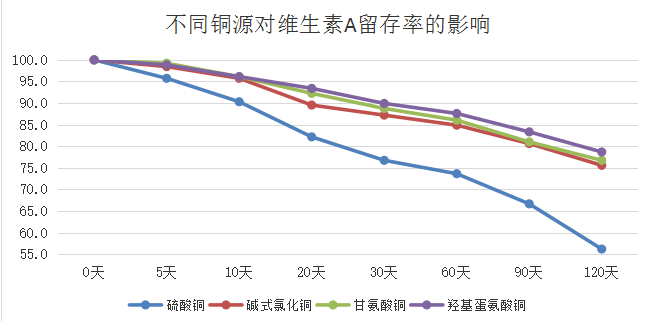
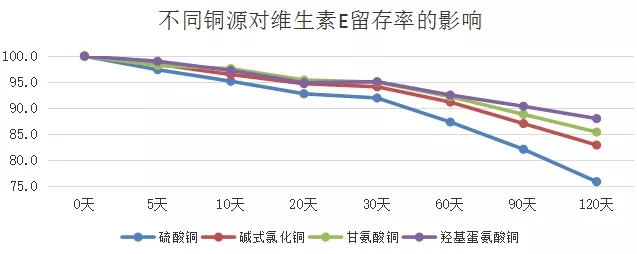
1.1 不同类型微量元素铜对维生素的破坏影响程度存在很大差异:有机微量元素对维生素的破坏程度相对较小,无机微量元素对维生素的破坏程度最严重的,羟基蛋氨酸系列氨基酸螯合物对维生素的破坏程度最小。
1.2 不同类型的维生素受微量元素铜的破坏程度存在一定差异:维生素E相对较稳定,受微量元素的影响程度相对较小,而维生素A受微量元素影响程度较大。维生素A影响水产动物的免疫与生长发育。
微量元素选择不合理会直接破坏饲料或预混料营养,影响饲料精准营养实现和稳定性,影响养殖效果。
2、微量元素铜的有效性
(1)溶失少
水产饲料,特别是虾蟹饲料因直接投放至水体中,被虾蟹抱食,摄食时间长,很容易造成可溶性养分的损失。因此,选择溶失率低、可利用的铜源,具有重要的的意义。
硫酸铜易溶于水,碱式氯化铜不溶于水,氨基酸络合铜,因工艺不同水溶性有一定差异,氨基酸螯合物不溶于水。
值得一提的是,所有氨基酸螯合微量元素如果未经过其他处理,均应该不溶于水。
(2)吸收高效性
相比无机硫酸铜,氨基酸络(螯)合铜,可以氨基酸通道吸收,少拮抗,吸收利用率高。
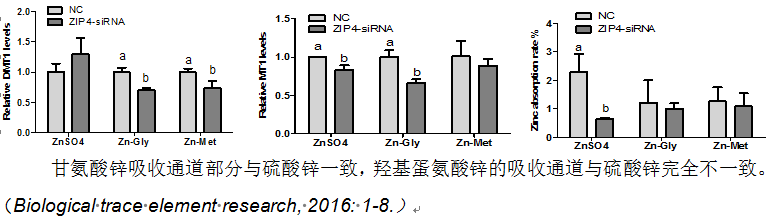
(3)体内可储存性,转运效率
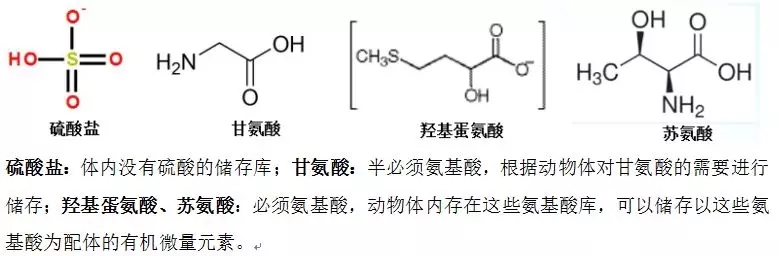
动物体内存在有效的特定氨基酸储存库 ,可以储存以特定氨基酸为配体的有机微量,在水产动物遭受免疫、环境应激时,可高效转运、及时调用,增强应激期间的抗应激和生产性能。
3、国家现有法规和政策变化
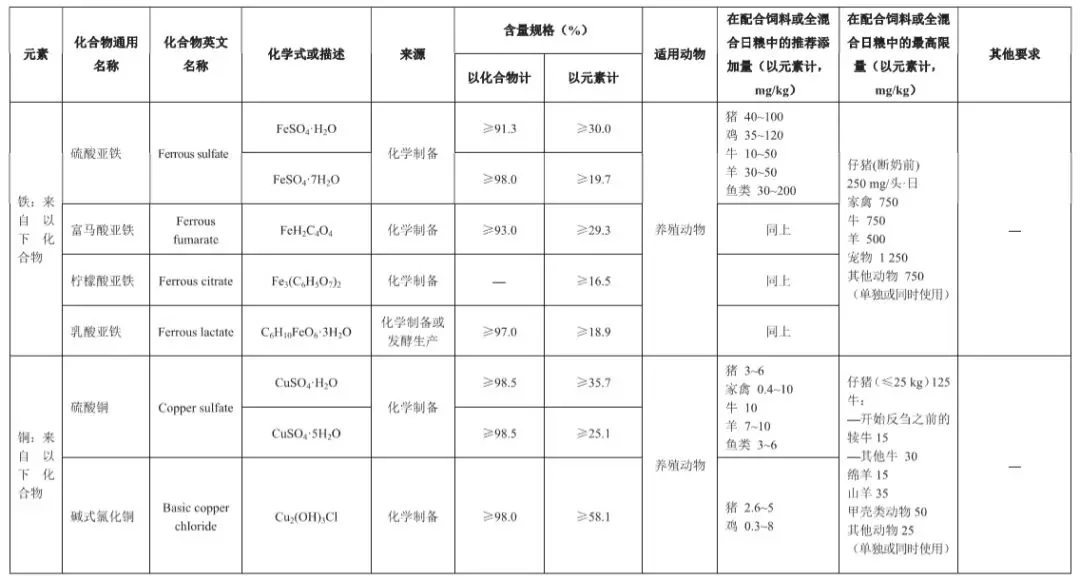
3.1 2625号法规规定:甲壳类动物最高限量50mg/kg饲料,其它水产动物25mg/kg饲料。
3.2 由于国家环保政策影响,湖泊、水库网箱越来越少,可用于饲料养殖生产的养殖水面越来越少,对于可能进一步限制或者严格要求养殖水体排放标准出台的可能性,认识和提前储备环保型微量元素铜应用技术是有必要的。
3.3 2020年饲料全面禁抗,认识和选择具有保健性功能营养的新型微量元素铜源,同样具有重要意义。
(2)市场常见铜源比较
表 不同铜源特点及代表产品

无机铜主要有硫酸铜,新型碱式氯化铜,被认为是一种更有效的无机铜源;有机铜的优势,一方面提现在更好的吸收储存,另一方面体现在更高效的调用利用性能;不同有机铜质量和结构差异较大,应用效果必然存在差异,应该注意有机铜源的选择。
(3)鱼饲料和虾饲料添加建议
随着后抗生素时代的到来,以及养殖集约化发展,环保要求越来越严格,消费者对高品质食品的需要,对养殖鱼虾微量元素营养铜添加量的建议,应该不止基于预防临床缺乏症或生长受损所需。以下建议是基于以上多个响应标准,和对水产动物生理、饲料特点及不同铜源的理解而设计。
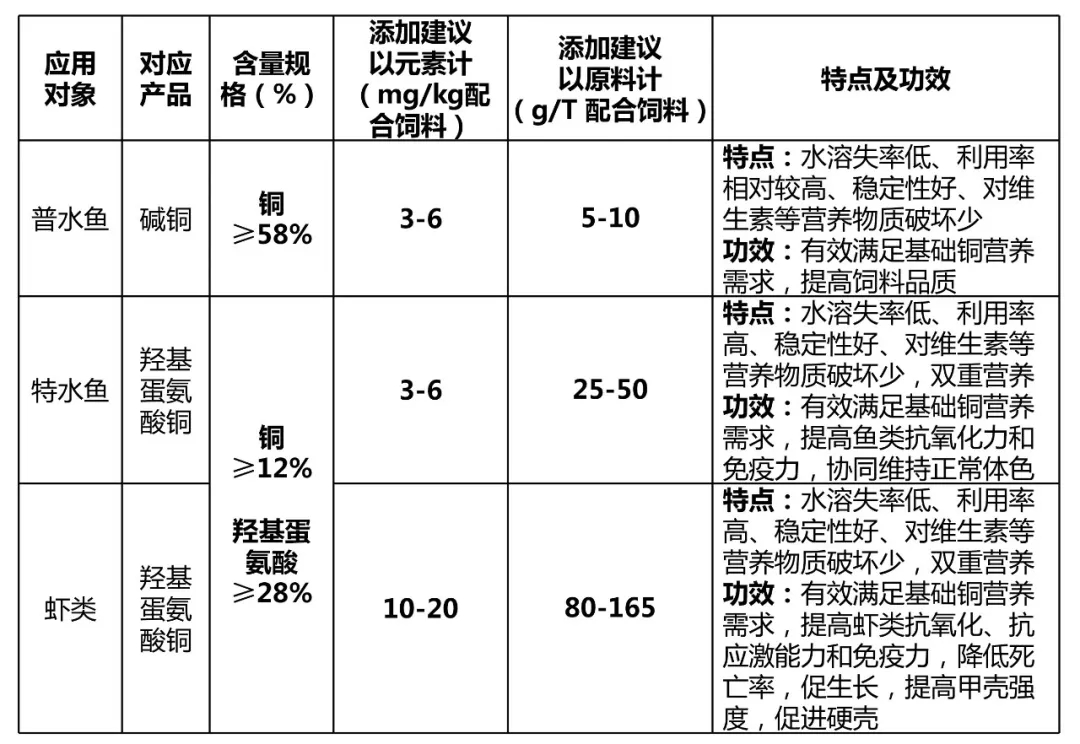
备注:蟹方面的铜营养研究非常少,其与虾同属甲壳类,生理特点也类似,建议添加量比对虾略高。无鳞鱼相比有鳞鱼,铜建议添加量可以略低。杂粕多,应激大,添加量越高。水体溶氧供应越差,养殖密度越高,添加量越大。
(4)羟基蛋氨酸应用案例
4.1 斜带石斑鱼幼鱼对羟基蛋氨酸铜的需求-----马豪勇等,2014年
表 饲料添加羟基蛋氨酸铜对斜带石斑鱼幼鱼生长和饲料利用,及对组织和血清非特异性免疫酶活性的影响,铜源为羟基蛋氨酸铜
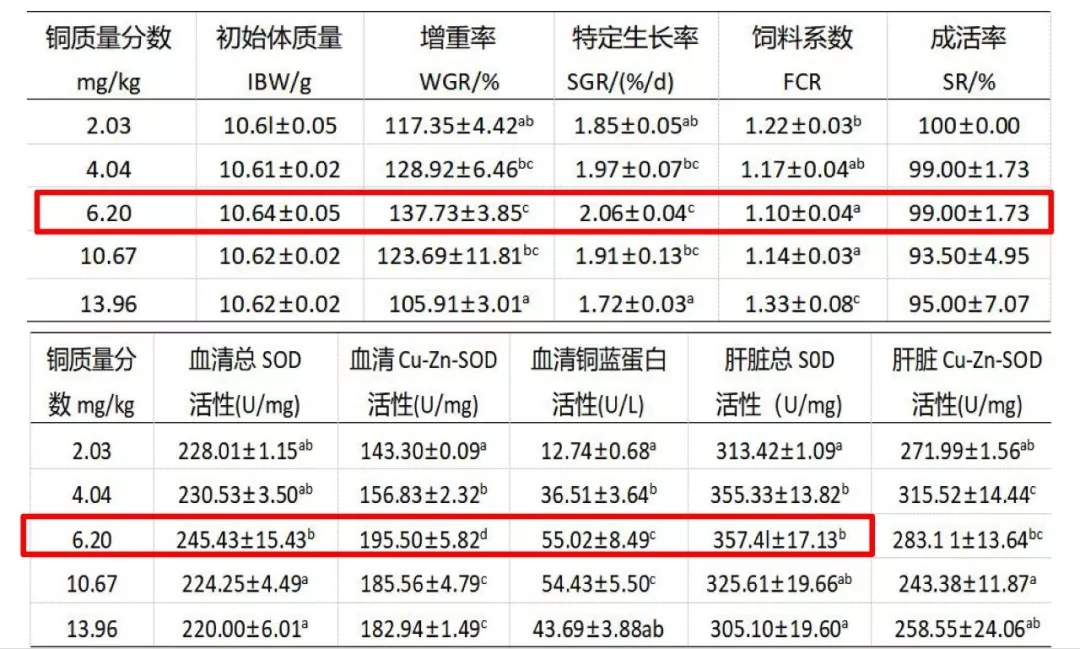
饲料中添加羟基蛋氨酸铜可促进斜带石斑鱼幼鱼生长提高饲料利用,同时能够提高部分非特异性免疫酶活性,增加机体组织铜含量。以特定生长率为判据,通过二次曲线模型拟合得出斜带石斑鱼幼鱼铜需要量为7.06mg/kg。
4.2 饲料中不同铜源对珍珠龙胆生长性能的影响-----殷彬,谭北平,2016年
表 不同铜源对珍珠龙胆生长性能的影响,5ppm指定铜源

相比无机硫酸铜,甘氨酸铜,羟基蛋氨酸螯合铜在提高增重率,特定生长率和饵料系数上效果最佳,增重率比硫酸铜组高33.4%,饵料系数降低3.1%。
4.3 日粮不同铜源和铜水平对异育银鲫生长性能、铜含量分布、血浆抗氧化活性和铜的相对生物利用率的影响-----邵仙萍,刘文斌等,2010年
表 不同铜源和铜水平对异育银鲫生长性能的影响
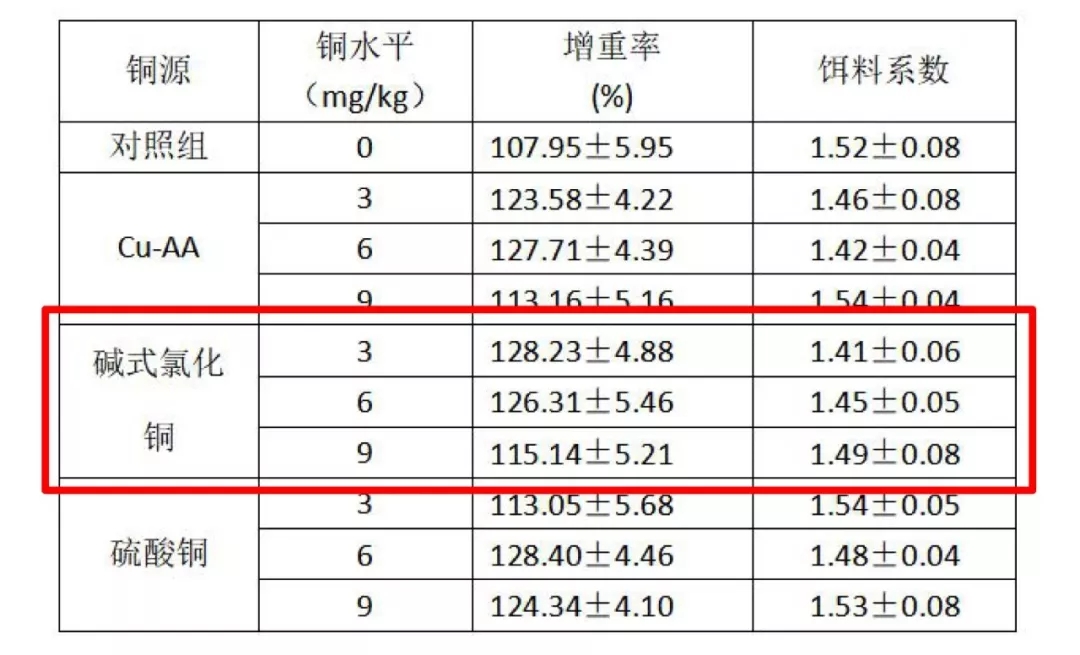
表 不同铜源的相对生物学利用率
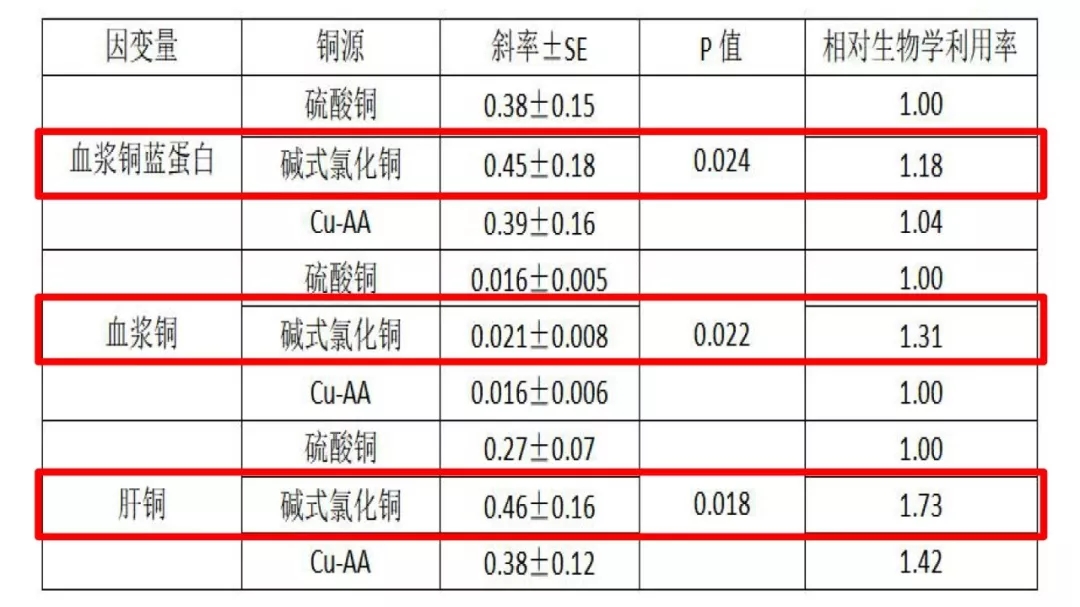
添加3-6mg/kg碱式氯化铜组增重率和饵料系数相对无机硫酸铜和复合氨基酸铜(蛋白水解盐)组更高。对肝铜、血浆铜、血浆铜蓝蛋白含量为评价指标,碱式氯化铜的相对生物学利用率分别比无机硫酸铜高73%、31%和18%,且均比复合氨基酸铜(蛋白水解盐)高。说明,在异育银鲫饲料中应用碱式氯化铜相对无机硫酸铜和复合氨基酸铜(蛋白水解盐)有更高的生物利用率。